

 Parlo adesso un po’ di questi nostri avi Cro-Magnon.
Parlo adesso un po’ di questi nostri avi Cro-Magnon.
Abbiamo reperti databili a poco più di 32.000 anni fa (tra i quali anche quelli riferibili all’Uluzziano (1) siciliano di Fontana Nuova, nelle campagne ragusane, dunque non tanto lontano da casa mia). I resti scoperti da Louis Lartet, un geologo francese, sarebbero poco posteriori, così come i molti affermano, a quelli dell’Uomo di Combe-Capelle (o Brünn, da non confondere assolutamente con quel Brünn Cro-magnoide che caratterizza le isole britanniche).
Ma anche sull’Uomo di Combe-Capelle potrei dire tanto, soffermandomi su questo caso specifico con un sunto molto importante. Hanno detto che sarebbe stato un ‘’antenato’’ del Cro-Magnon, essendo ciò erroneo, semmai il contrario: trattasi di un’evoluzione del Cro-Magnon in una forma più armonica, leptoprosopa e non euridolicormorfa considerata ‘’disarmonica’’ (evidente ancora nella varietà Oberkassel, con la quale proprio il Brünn si incontrava, creando così altre varietà centro-europee e Nord-occidentali). Ma ciò che più è da ritenersi erroneo è lo sbrigativo corollario sulla sua provenienza e descrizione, che quanto mai risulta essere del tutto fuorviante all’evidenza stessa. Questo tipo umano sarebbe giunto in Europa dall’Asia ed i resti più antichi risalirebbero a 42.000 anni fa, ritrovati essi nell’omonima località francese, dove questo tipo umano avrebbe condiviso questo ambiente con i più vecchi Neandertaliani. Ma lo scheletro ritrovato nel 1909 a Monferrand ed associato a questo tipo umano, in base alle analisi condotte sugli isotopi carbonici, deve essere datato al Mesolitico, intorno al 7.600-7.500 a.C. Questo tipo umano presenta a detta loro caratteristiche ‘’paleo-mediterranee’’ (come già detto, trattasi semplicemente di una forma leptoprosopa e più armonica del Cro-Magnon più arcaico), imparentata a sua volta al tipo paleo-australoide, ovvero gli aborigeni australiani (alla razza veddide, cosa assolutamente da escludere), se non addirittura capostipite dell’uno e dell’altro (ipotesi da rigettare totalmente, assolutamente infondata). La volta cranica si presenta alta, pentagonoide, circa 1.400 cm³, con fronte bassa, sfuggente (quando è invece leggermente inclinata, proprio come la nostra che da essa deriva infatti); arcate sopracciliari prominenti e orbite basse; leptoprosopia, camerrinia, e prognatismo; statura medio-bassa (tutti dati assolutamente sbagliati, basta vedere la ricostruzione dello splacnocranio). A questo tipo vengono ascritti i ritrovamenti ossei dei due esemplari rinvenuti in Moravia, a Brno, risalenti al 25.000 a.C.; quelli di due esemplari russi, di Pavlov, risalenti al 25.000 a.C.; quelli di un giovane maschio ritrovati in Liguria, nelle Arene Candide, e risalenti al 18.000 a.C.; e quelli di giovane maschio ritrovati in Puglia, nella Grotta Paglicci del Gargano, e risalente al 18.000 a.C. Il fatto è che i resti dei due giovani maschi rinvenuti in Italia, Liguria e Puglia, appartengono in realtà al tipo Cro-Magnon più arcaico; e tutti questi ritrovamenti, dalla Russia all’Italia e alla Francia, sono stati fatti in contesti dell’orizzonte culturale gravettiano, dunque trattasi di Cro-Magnon più vicini alla varietà Oberkassel. Gli Ainu dell’isola di Hokkaidō, dei quali si è ipotizzata l’ascendenza dal tipo Combe-Capelle presentano invece caratteristiche diverse, pur sempre non distanti a quelle del Cro-Magnon più arcaico, e lo si vede dalla fronte dritta, e questo tratto fisiognomico è tipico dell’Homo Europaeus, ossia il Cro-Magnon.
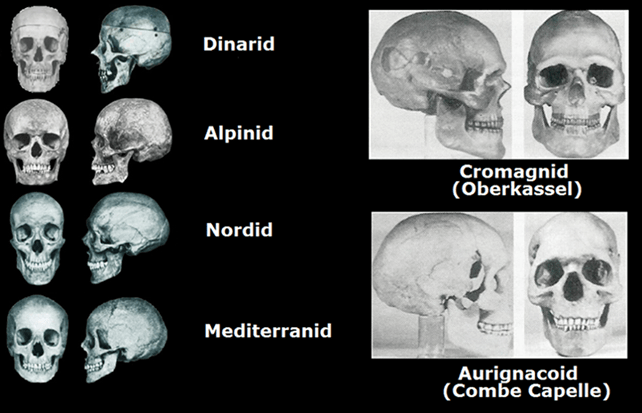
Le caratteristiche del Cro-Magnon, facendola molto semplice e breve, sono le seguenti: dolicomorfia cranica (lunghezza considerata dalla fronte all’occipite); grande capacità della volta cranica, fino a 1650 cm³; cavità orbitali rettangolari e basse; naso prominente con glabella alta; ortognatismo (volto dritto) e squadrato; alta statura media, intorno a 1,80 m. per i maschi (a volte oltre 1,90 m.), fino a rari casi di 1,68-1,70 m. per le femmine (anche 1,90 m., come nel caso del primo ritrovamento dei Balzi Rossi in Liguria). Un particolare, rispetto alla varietà aurignaziana, è l’andamento sagittale che passa tra la sutura dei parietali fino alla sutura occipitale e poi a scendere: nei Cro-Magnon l’andamento lineare sagittale sembra al tocco (potete farlo anche voi a casa, passando un dito della mano) quasi rettilinea, per poi toccare un avvallamento (una schiacciatura) a 45°, e poi una sommità (come un bernoccolo) e poi di nuovo a scendere verso la nuca; nella variante aurignaziana questa linea sagittale sembra al tocco un po’ ondulata, passando tra i due parietali in modo progressivo, a salire (leggera ipsicefalia, cranio all’insù), e poi, compiendo lo stesso percorso scendendo verso la nuca, noterete sempre questa ondulazione, come se gli angoli descritti fossero smussati (nei Cro-Magnon una regolare spigolosità, che non c’è più nel Capelloide).
Insomma i nostri avi Cro-Magnon sono (o meglio dire erano?) il tipo nordide, detto anche Homo Europaeus. Le cladi mitocondriali dominati eurasiatiche (dalla Siberia a noi occidentali) erano U2, U5 e U8 (il cosiddetto DNA mitocondriale di ‘’Ursula’’, secondo l’ironia di Bryan Sykes nel suo saggio The Seven Daughters of Eve, pubblicato nel 2001), stando a quanto dice la gran maggioranza degli studiosi che recentemente se ne è occupata(2) . Vi sono poi le varianti occidentali: X, T e H che hanno marcato il Paleolitico; poi K e V nel corso del Mesolitico; ed infine a seguire tutti quelli neolitici giunti sino alla nostra epoca. Secondo il team di Bramanti, i resti fossili di un individuo aurignaziano rinvenuto nella grotta di Fumane nel Veneto hanno fornito gli stessi dati genotipici dei resti rinvenuti a Ust’ Išim, in quel di Siberia. Purtroppo gli aplogruppi individuabili dal cromosoma Y sono i più difficili da trattare, in quanto ce l’hanno solo i maschi, è soltanto uno e con possibilità di destrutturazione nel corso temporale. Dai resti di un bambino d’età tra i 3 ed i 4 anni rinvenuti in Siberia ed appartenenti alla facies archeologica di Mal’ta-Buret’, si è evinto l’aplogruppo Y-DNA R, ossia l’antenato di R1, dal quale a sua volta derivano R1a e R1b, ed R2, ovvero quelli nostri, attuali. Dalla penisola iberica alla Francia (si pensi alle pitture rupestri di Lascaux), da lì alla Scandinavia, da lì ancora alla grotta di Uluzzo nel leccese e nel territorio di Ragusa in Sicilia (si pensi anche all’arte figurativa dell’isola di Levanzo ed a quella della grotta dell’Addaura, nel palermitano), e dunque fino alla Siberia, vi è lo stesso tipo (genotipo e fenotipo) umano, il Cro-Magnon. Il genoma autosomico delle attuali popolazioni europee (non parlo degli allogeni-alloctoni ovviamente) deriva dai cacciatori-raccoglitori paleo-mesolitici, dagli agricoltori neolitici e da una piccola parte di Siberiani del Paleolitico. Alcuni studiosi ritengono in base ai risultati delle loro ricerche che gli attuali abitanti dei paesi che si affacciano sul Mar Baltico hanno maggior DNA autosomico correlabile ai cacciatori-raccoglitori paleo-mesolitici europei (3) . Ed altra cosa da sfatare è che questi nostri avi non vivevano nelle grotte, ma nelle capanne. Le grotte, gli ambienti rupestri o ipogei erano luoghi di culto. La diffusione dei Cro-Magnon nel continente eurasiatico è individuabile dalla sovrapposizione della Cultura castelperroniana su quella musteriana del tempo dell’Homo Neanderthalensis, dunque a partire da 40.000 anni fa.
Ad esempio, proprio quando ho parlato dei Popoli del Mare, gli Haunebu, nei quali figuravano anche i non ‘’Siculi’’ Shekelesh, ho menzionato i Libues biondi, quelli che hanno lasciato memoria nel coronimo dell’attuale Libia. Ma non erano i soli ad essere alti, biondi, rossi e con gli occhi chiari in quella regione. Si pensi anche ai Guanci delle isole Canarie, popolo che stupì gli Spagnoli ai tempi delle grandi conquiste oltre l’oceano. Ancora oggi, alcuni Berberi presentano queste peculiarità fenotipiche (dunque ed in primis genotipiche). Il tipo cro-magnoide di cui sto parlando è infatti quello di Mechta-Afalou. Si pensa che i Guanci siano stati discendenti dei Berberi, ma non è proprio così. A parte il fatto che oggi come oggi non tutti i Berberi presentano le caratteristiche originarie (l’ecumene islamica ha fatto gli stessi danni del cristianesimo), anche loro allo stato attuale si presentano molto ibridati (un vero peccato); ma poi questi Berberi, quelli più antichi, discenderebbero dai Libues biondi (vi sono tracce della loro biondezza persino sulle pitture rupestri sahariane), a sua volta discendenti da quel Cro-Magnon che ci ha lasciato in eredità anche i Guanci, ormai purtroppo estinti. In ogni caso, la popolazione nordica della Cultura maglemosiana (4) (i dalo-falici), quella delle Culture del Dneper-Donets e Sredny-Stog (gli addomesticatori di cavalli), quella delle Culture siberiane di Afanasevo e Tagar, quella giunta fino all’estremità dell’Asia ed in Nord America (Homo di Kennewick, ben prima delle altre popolazioni volgarmente dette ‘’di pelle rossa’’), quella pirenaica delle celebri pitture e via dicendo, sono tutte discendenti, figlie di un unico padre, nostro progenitore Cro-Magnon.
Alessandro Daudeferd Bonfanti
Note
1) Dal nome della Grotta Uluzzo, nel territorio di Lecce, Puglia. Ed il caso di Fontana Nuova di Ragusa non è Aurignaziano pieno, come solito si legge, ma una fase precedente, coincidente con la fine del Castelperroniano, ossia l’Uluzziano appunto.
2) S. Benazzi, V. Slon & S. Talamo, The makers of the Protoaurignacian and implications for Neandertal extinction, in Science, Vol. 348, n. 6236, pagg. 793-796; B. Bramanti, M.G. Thomas & W. Haak, Genetic Discontinuity Between Local Hunter-Gatherers and Central Europe’s First Farmers, in Science, Vol. 36, n. 5949, pagg. 137-140; Q. Fu, A. Mittnik & P.L.F. Johnson, A Revised Timescale for Human Evolution Based on Ancient Mitochondrial Genomes, in Current Biology, Vol. 23, n. 74, pagg. 553-559.
3) I. Lazaridis, N. Patterson & A. Mittnik, Ancient human genomes suggest three ancestral populations for present-day Europeans, in Nature, Vol. 513, n. 7518, pagg. 409-413.
4) Non dimenticate mai che dalla Cultura di Maglemose deriva quella di Ertebølle (villaggio della Danimarca), e da questa ancora la Cultura del vaso imbutiforme (Trichterbecherkultur dal 4.000 al 2.700 a.C. circa), così come le due Culture sovrappostesi della ceramica cordata/ascia da combattimento e dell’anfora globulare (dal 3.200 fino al 1.800 a.C., quindi durante il tardo neolitico, il calcolitico e la prima età del Bronzo), e ancora quella che ha interessato anche la Sicilia nel corso del III millennio a.C. con i noti dolmen e la tipica cultura materiale, ossia la Cultura del bicchiere campaniforme, diffusasi dal Centro-Nord Europa a partire dal 2.900/2.800 fino al 1.800 a.C.














